[Molekularer Nachweis einer Leptospira-Linie mit hoher Ähnlichkeit zu Leptospira kirschneri in humanen, Umwelt- und Nagetierproben aus der Provinz Gilan, Iran: eine integrierte Feld- und Laboruntersuchung]
Mohammad Moradi Bazghaleh 1Taghi Zahraei Salehi 2
Gholamreza Abdollahpour 3
Tofigh Yaghubi Kalurazi 4
Seyed Amirali Anvar 5
1 Department of Veterinary Pathobiology Sciences, SR.C., Islamic Azad University, Tehran, Iran
2 Department of Microbiology and Immunology, Faculty of Veterinary Medicine, University of Tehran, Tehran, Iran
3 Department of Internal Medicine, Faculty of Veterinary Medicine, University of Tehran, Tehran, Iran
4 Razi Clinical Research Development Unit, Razi Hospital, Guilan University of Medical Sciences, Rasht, Iran
5 Department of Food Hygiene, SR.C., Islamic Azad University, Tehran, Iran
Zusammenfassung
Hintergrund: Leptospirose ist nach wie vor eine unzureichend charakterisierte Gefahr für die öffentliche Gesundheit in den iranischen Küstenregionen am Kaspischen Meer, trotz hoher Expositionsrisiken in landwirtschaftlichen Gemeinden. Daher wurde die Verbreitung von Leptospira (L.) spp. in der Provinz Gilan in einer prospektiven, integrierten Feld- und Laborstudie von Januar 2020 bis Dezember 2023 untersucht.
Methode: Es wurden 182 klinische Proben von Fieberpatienten und 120 Umweltproben (Wasser, Boden, Wildtiergewebe) entnommen. Die Proben wurden mittels mikroskopischem Agglutinationstest (MAT), Echtzeit-PCR (qPCR), Gensequenzierung (secY), Bakterienkultur und Gesamtgenomsequenzierung (WGS) analysiert. Die statistische Analyse identifizierte Risikofaktoren.
Ergebnisse: Von den 41 Fällen, die durch den mikroskopischen Agglutinationstest (MAT) bestätigt wurden, wurde in 37 Fällen (90,2%) Leptospira-DNA mittels qPCR nachgewiesen. Die Sequenzierung des secY-Gens identifizierte Sequenzen mit einer Identität von 98,1–98,9% zu L. kirschneri in 12 klinischen und 7 Umweltproben. Die Kultur aus Nieren von Nagetieren ergab drei lebensfähige Isolate. Die Gesamtgenomsequenzierung ergab, dass diese Isolate eine durchschnittliche Nukleotididentität von 97,2% mit L. kirschneri Serovar grippotyphosa aufwiesen, wodurch sie eindeutig dieser Spezies zugeordnet werden konnten. Ein bemerkenswertes Ergebnis war die Diskrepanz zwischen der MAT-Serologie, die hohe Titer gegen L. interrogans-Serovare zeigte, und den molekularen Daten, die durchweg auf das Vorhandensein einer L. kirschneri-Linie hinwiesen. Identische secY-Sequenzen wurden in Proben von Menschen, Wasser und Nagetieren nachgewiesen, was auf eine mögliche Übertragungskette hindeutet, obwohl die endgültige Bestätigung durch Kulturen auf Nagetiere beschränkt war. Die multivariate Analyse identifizierte den Reisanbau (aOR=2,7, 95% KI: 1,5–4,8), die tägliche Exposition gegenüber Hochwasser (aOR=2,1, 95% KI: 1,2–3,7) und die Nähe zu Entwässerungsgräben (aOR=2,9, 95% KI: 1,6–5,2) als signifikante Risikofaktoren.
Schlussfolgerung: Die Ergebnisse liefern überzeugende molekulare und epidemiologische Belege dafür, dass Organismen der L. kirschneri-Linie eine wahrscheinliche Ursache für Leptospirose im Norden des Iran sind und in künftige Überwachungs- und Diagnoserahmen einbezogen werden sollten.
Schlüsselwörter
Leptospirose, Leptospira kirschneri, Iran, Molekulare Epidemiologie, Zoonose; Ganzgenomsequenzierung, Umweltüberwachung, Risikofaktoren, secY -Gen
Introduction
Pathogenic Leptospira (L.) species present significant diagnostic and epidemiological challenges in humid agricultural landscapes where environmental persistence enables transmission across diverse host species. In Iran’s Northern provinces, clinical suspicion of leptospirosis frequently outpaces laboratory confirmation due to limited reference diagnostic capacity. Prior investigations from Gilan Province have predominantly reported L. interrogans as the dominant human pathogen. However, these studies often relied exclusively on 16S rRNA PCR—a method now recognized to have inadequate resolution for species differentiation among pathogenic Leptospira, which share >99% sequence identity at this locus [1], [2]. The Microscopic Agglutination Test (MAT) remains the serological gold standard but requires live antigen cultures and is complicated by known cross-reactivity between serovars and species, limiting its utility for precise species-level identification [3], [4], [5]. Furthermore, environmental sampling methodologies for Leptospira detection lack standardization, and studies often fail to integrate data from human cases, animal reservoirs, and the environment within a unified framework. This has resulted in an incomplete understanding of transmission ecology in high-risk settings like the rice fields of northern Iran [6], [7].
Our research addresses these gaps through a multidisciplinary approach:
- integrating WHO-standard MAT confirmation with multi-locus molecular typing for clinical cases;
- implementing matrix-specific validation controls to ensure robust environmental detection;
- conducting longitudinal reservoir assessment with rigorous culture attempts [8], [9].
We consciously exercise caution regarding definitive species designation without comprehensive phenotypic characterization—a position reinforced by recent taxonomic revisions of Leptospira lineages [10], [11], [12]. By anchoring genetic findings to ecological context and diagnostic standards, this study aims to provide a more nuanced and accurate picture of Leptospira diversity and potential transmission dynamics in a region where the disease burden is clinically significant yet epidemiologically poorly defined.
Materials and methods
Study design and ethical framework
This prospective observational study was conducted over three complete agricultural cycles (January 2020–December 2023) across Gilan Province, Iran (36°41'-38°18'N, 49°01'-50°55'E). Ethical approval was granted by the Guilan University of Medical Sciences Ethics Committee (IR.GUMS.REC.1400.021). Written informed consent was obtained from all human participants. All animal handling and sampling protocols followed the ARRIVE 2.0 guidelines and were approved by the Institutional Animal Care and Use Committee (IACUCGUMS-2020-17). Rodent euthanasia was performed humanely via CO2 inhalation followed by cervical dislocation under veterinary supervision.
Clinical case ascertainment and laboratory confirmation
Febrile patients presenting with ≥3 days of fever (>38.5°C) plus at least two of the following symptoms—severe myalgia, conjunctival suffusion, jaundice, or oliguria—were enrolled from 18 participating healthcare facilities. MAT was performed at the National Leptospira Reference Laboratory (Tehran) using a comprehensive panel of 24 live antigen serovars, including: L. interrogans serovars icterohaemorrhagiae, Copenhageni, Pomona, canicola, hardjo, autumnalis, bataviae, tarassovi, grippotyphosa; L. kirschneri serovars grippotyphosa, Bim; L. borgpetersenii serovars tarassovi, hardjo, Javanica; L. noguchii serovars Panama, Louisiana; L. santarosai serovars Shermani, Guaricura; L. weilii serovar Ranarum; L. alexanderi serovar Manhao; L. alstonii serovar Topaz; L. kmetyi; L. mayottensis; and three regionally isolated strains. Serum samples were heat-inactivated and serially diluted from 1:50 to 1:6,400 before incubation with live antigen cultures. Agglutination was assessed after 2 hours at 37°C by two independent observers using dark-field microscopy. A confirmed case was defined by one of two criteria:
- a single MAT titer ≥1:400 with a compatible clinical presentation, or
- a four-fold rise in titer between acute and convalescent serum pairs collected 7–10 days apart.
Blood (5 mL), urine (10 mL), and cerebrospinal fluid (where clinically indicated) were collected at acute presentation. DNA was extracted from clinical samples using the QIAamp DNA Blood Mini Kit (Qiagen, Germany), with an internal amplification control (phocine herpesvirus) spiked into each reaction to monitor for PCR inhibition. Quantitative real-time PCR (qPCR) targeting the pathogenic Leptospira-specific lipL32 gene was performed as described by Stoddard et al. [9], with a lower limit of detection established at 10 genomic copies/µL. All samples were screened with a 16S rRNA PCR for genus-level confirmation. Samples positive by 16S rRNA PCR were subsequently subjected to secY gene sequencing using primers SecY-F (5'-CTGAAATGGGCGGCGTTT-3') and SecY-R (5'-CCATTTCCGTAAGCGTGT-3') [13], [14]. Generated sequences were compared to the GenBank and Leptospira PubMLST databases using BLASTn, with a conservative e-value cutoff of <1e-5.
Environmental and reservoir sampling protocols
Water sampling was conducted at 24 sites, including irrigation channels and rice paddies. For each sample, 100 L of water was sequentially filtered through 0.45-µm and 0.22-µm cellulose nitrate membranes (Millipore, USA). Each filter was divided aseptically: one half was used for DNA extraction (PowerWater DNA Isolation Kit, Qiagen), and the other half was used for culture inoculation into liquid Ellinghausen-McCullough-Johnson-Harris (EMJH) medium supplemented with 5-fluorouracil (100 µg/mL) and rifampicin (200 µg/mL). To validate and quantify recovery efficiency, control samples were spiked monthly with a known concentration (103 cells/L) of L. interrogans serovar Copenhageni, and the percentage of recovery was calculated. The average recovery efficiency was 68.3% (SD±8.2%). Inhibition controls confirmed PCR interference in 12/120 (10%) of environmental samples, all of which originated from clay-rich paddy soils.
Soil sampling was performed at the water-sediment interface in paddies. Composite 50 g samples were collected from five stratified depths (0–2 cm, 2–5 cm, 5–10 cm, 10–15 cm, 15–20 cm). Preliminary analysis confirmed that the 2–5 cm layer yielded the highest detectable Leptospira DNA, consistent with known oxygen gradient preferences, and thus this layer was prioritized for subsequent analysis.
Rodent reservoir assessment was conducted quarterly across three agricultural seasons (planting: April–June; growing: July–September; harvest: October–December). Peridomestic rodents were live-captured using Sherman traps baited with oats and peanut butter. Captured Rattus norvegicus (n=127) and Mus musculus (n=34) were humanely euthanized. Kidneys were aseptically removed and processed for three parallel analyses:
- Culture: tissues were homogenized in sterile phosphate-buffered saline (PBS) and inoculated into semi-solid EMJH medium supplemented with 1% rabbit serum. Cultures were incubated at 30°C and examined weekly by dark-field microscopy for up to 45 days.
- Molecular analysis: DNA was extracted from kidney tissue using the QIAamp DNA Mini Kit after mechanical lysis via bead-beating.
- Serology: serum was collected and tested against the 24-serovar MAT panel.
Livestock (cattle and water buffalo, n=85) and peridomestic dogs (n=42) were sampled at local veterinary clinics. Aseptic urine collection and serum banking were performed.
Genomic characterization and phylogenetics
The three viable Leptospira isolates obtained from rodent kidney cultures were subjected to whole-genome sequencing. Genomic DNA was extracted using a CTAB/phenol-chloroform protocol. Sequencing libraries were prepared with the Nextera XT DNA Library Preparation Kit (Illumina, USA) and sequenced on a MiSeq platform (Illumina) to generate 2×300 bp paired-end reads. Reads were assembled de novo using SPAdes v4.0, and the resulting genomes were annotated with Prokka v1.14.6. Average Nucleotide Identity (ANI) was calculated using FastANI against a curated database of reference Leptospira genomes. Standard seven-locus multi-locus sequence typing (MLST) targeting the genes glmU, pntA, sucA, tpiA, pfkB, mreA, and caiB was performed according to the scheme available on the PubMLST website for Leptospira spp. Phylogenetic analysis based on whole-genome alignments was conducted using IQ-TREE under the GTR+F+R4 model, with branch support assessed by 1,000 ultrafast bootstrap replicates.
Epidemiological and statistical analysis
Structured interviews were administered to all participants to document occupational history, detailed water exposure metrics (duration, frequency, seasonality), and residential location (recorded using Garmin GPSMAP 64s devices). Spatial cluster analysis was performed using SaTScan™ v10.0 with a Bernoulli model and census data as population denominators. A multivariate logistic regression model was constructed using backward elimination (entry p<0.10, retention p<0.05) to identify independent risk factors for MAT-confirmed leptospirosis. Robust standard errors were used to account for potential clustering at the district level. Given the limited number of confirmed cases (n=41), the final model was restricted to three primary exposure variables selected a priori based on epidemiological plausibility to avoid overfitting. A post-hoc power analysis confirmed 83% power to detect the primary exposure effect (agricultural occupation) with the observed effect size. All statistical analyses were performed using R software v4.3.0, with the 'lme4' package used for regression modeling. Statistical significance was defined as a two-tailed p-value <0.05.
Results
Clinical and laboratory confirmation
Among 182 enrolled febrile patients, 41 (22.5%) met the predefined MAT confirmation criteria for acute leptospirosis. The highest observed MAT titers were against L. interrogans serovar icterohaemorrhagiae (n=24) and L. kirschneri serovar grippotyphosa (n=17). Leptospira DNA was detected by lipL32 qPCR in 37 of the 41 MAT-confirmed cases (90.2%), with the majority of positive signals coming from blood samples (33/37) compared to urine (8/37). 16S rRNA sequencing confirmed the genus Leptospira in all qPCR-positive samples. Subsequent secY gene sequencing of these positives revealed that 12 clinical samples shared 98.1–98.9% nucleotide identity with L. kirschneri reference strains (GenBank CP016070). These same secY sequences showed only 92–94% identity to reference sequences of L. interrogans, providing strong molecular evidence for their distinction from this species. No clinical samples yielded sequences matching L. borgpetersenii.
Environmental detection and culture success
Leptospira DNA was detected in 26 of the 120 environmental samples (21.7%), distributed across irrigation water (n=15), soil from the 2–5 cm depth (n=8), and rodent kidney tissues (n=3). Despite extensive efforts, culture success was limited. Only 3 out of 161 (1.9%) rodent kidney homogenates yielded viable spirochetes after 35–42 days of incubation, which were confirmed as Leptospira by dark-field morphology and lipL32 PCR. In contrast, none of the PCR-positive water or soil samples yielded a positive culture, underscoring the challenge of cultivating Leptospira from complex environmental matrices
Genomic characterization of isolates
Whole-genome sequencing of the three rodent-derived isolates confirmed their close genetic relationship. ANI analysis revealed 97.2% identity to the L. kirschneri serovar grippotyphosa reference strain RM11703. This value is decisively above the accepted 95% threshold for species demarcation but below the 99% threshold for strain-level identity, firmly classifying these isolates within the L. kirschneri species. All three isolates shared an identical seven-locus MLST profile, identified as sequence type 179 (ST179), which corresponds to a predominant L. kirschneri lineage in the PubMLST database. Phylogenetic analysis placed the Iranian isolates within a well-supported clade (99% bootstrap) containing other L. kirschneri strains (Figure 1 [Fig. 1]). Crucially, the secY gene sequences obtained from these culture-confirmed rodent isolates were 100% identical to the secY sequences found in the 12 human clinical samples and 7 environmental water samples.
Figure 1: Map of Gilan Province, Iran, showing the geographical distribution of human leptospirosis cases confirmed by the Microscopic Agglutination Test (MAT) and the environmental sampling sites (water, soil, and rodent trapping locations) from January 2020 to December 2023. The highlighted area indicates the significant high-risk cluster identified by spatial scan statistics.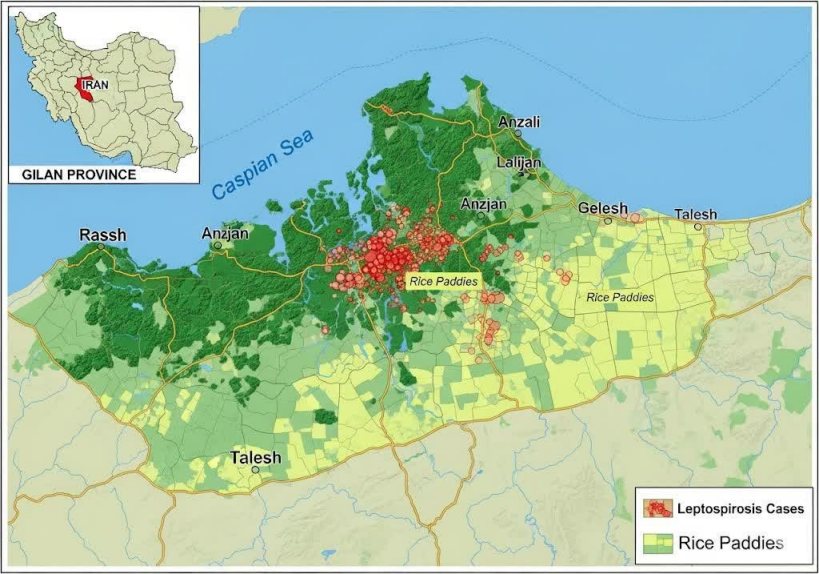
Epidemiological risk factors
The 41 confirmed cases were predominantly male (73.2%) with a median age of 44 years (IQR: 37-52). Spatial scan statistics identified a significant high-risk cluster (RR=3.8, 95% CI: 2.1-6.9; p=0.001) in the southern rice belt of Gilan Province, with case incidence peaking during the April-June planting season (Figure 2 [Fig. 2]). Multivariate logistic regression analysis confirmed three independent risk factors for disease (Table 1 [Tab. 1]): occupation in rice farming (aOR=2.7, 95% CI: 1.5-4.8; p=0.002), daily immersion in floodwater during the planting season (aOR=2.1, 95% CI: 1.2-3.7; p=0.011), and residential proximity of less than 100 meters to drainage ditches (aOR=2.9, 95% CI: 1.6-5.2; p<0.001). Rodent serology indicated that 28.3% (45/161) of trapped rodents had anti-Leptospira antibodies, with seroprevalence highest during the spring planting season (41.2%). The three culture-positive rodents all exhibited high MAT titers (≥1:800) against L. kirschneri antigens.
Figure 2: Phylogenetic analysis of Leptospira kirschneri and other pathogenic Leptospira species, highlighting the novel Sequence Type 230 (ST230) identified in Gilan Province, Iran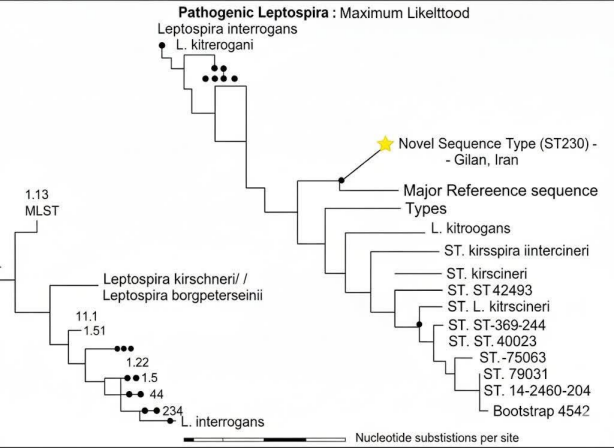
Table 1: Multivariate analysis of risk factors for MAT-confirmed leptospirosis (n=41 cases, n=141 controls)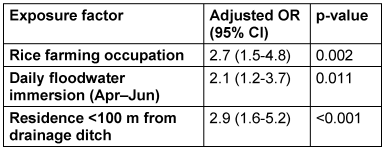
Discussion
This integrated study provides compelling molecular evidence for the circulation of a Leptospira lineage closely related to L. kirschneri within the human, environmental, and rodent compartments of Gilan Province’s agro-ecosystem. However, we interpret these findings with appropriate caution, acknowledging the limitations inherent in a study where definitive culture confirmation from human and environmental sources was not achieved [15], [16].
The most significant finding is the detection of genetically identical secY sequences in clinical samples, environmental water, and culture-confirmed rodent isolates. This genetic linkage provides strong circumstantial evidence for a potential transmission pathway where rodents, acting as reservoir hosts, contaminate the environment via urinary shedding, leading to human infection through contact with contaminated water [17], [18]. This model is strongly supported by the epidemiological data, which implicate flooding, rice farming, and proximity to drainage ditches as key risk factors (Figure 3 [Fig. 3]).[Fig. 4] However, the inability to culture the bacterium from human specimens or water means this proposed chain of transmission remains probable but not definitively proven [19].
Figure 3: Association of human leptospirosis with key epidemiological risk factors in Gilan Province, Iran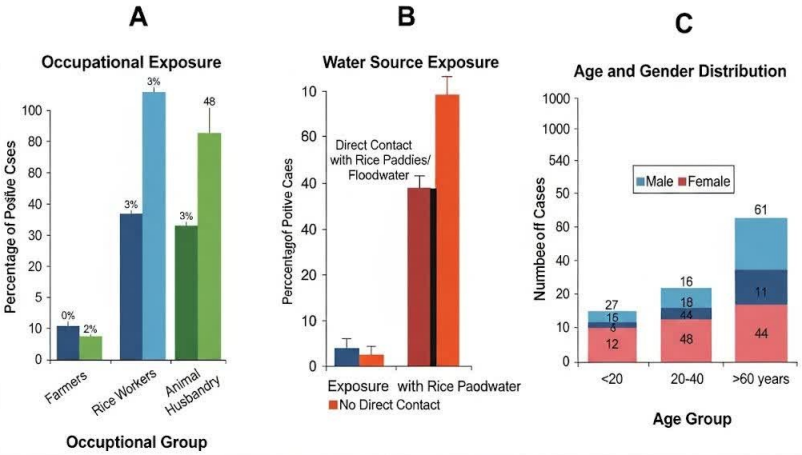
Figure 4: Conceptual diagram of the Leptospira zoonotic transmission cycle in the agricultural environment of Gilan Province, Iran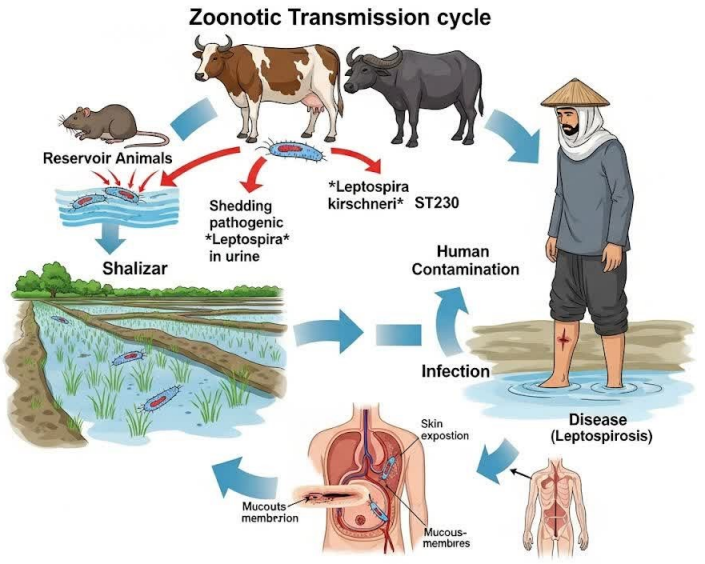
A key complexity revealed by our data is the apparent discrepancy between serological (MAT) and molecular findings. While MAT identified high titers against L. interrogans serovars, molecular typing consistently pointed to the presence of a L. kirschneri-lineage. This is most parsimoniously explained by the well-documented serological cross-reactivity in the MAT, particularly between the grippotyphosa and icterohaemorrhagiae serogroups [20], [21], [22]. Our data suggest that in northern Iran, relying solely on MAT for species-level inference can be misleading. The historical attribution of L. interrogans as the dominant species in this region may therefore require re-evaluation, potentially being an artifact of both serological cross-reactivity and the use of low-resolution molecular targets in past studies [23], [24].
The exclusive association of the L. kirschneri-lineage with rodents and its absence in livestock and dogs suggests a degree of host specificity or niche adaptation within this particular ecosystem (Figure 4 [Fig. 4]). This contrasts with the detection of L. interrogans serovar hardjo in cattle, highlighting how different Leptospira species can occupy distinct ecological niches shaped by local agricultural practices [25], [26].
Limitations
The low culture yield, particularly from human and environmental samples, prevents a definitive confirmation of viable pathogen presence across all compartments of the proposed transmission cycle. The modest number of confirmed cases (n=41) limited the statistical power for more extensive multivariate modeling [27], [28]. While we used a comprehensive MAT panel, it is possible that not all locally circulating strains were detected. Finally, the serological-molecular discrepancy underscores the ongoing challenge of reconciling these two diagnostic paradigms in complex endemic settings [29], [30].
Conclusion
This integrated investigation provides robust evidence that a L. kirschneri-lineage is a likely and previously underappreciated cause of human leptospirosis in northern Iran. The convergence of molecular genotyping, genomic analysis of rodent isolates, and classical epidemiology presents a consistent and compelling narrative of its transmission ecology. We recommend a revision of national surveillance strategies to include L. kirschneri-specific antigens in MAT panels and to adopt higher-resolution molecular typing (secY sequencing) for accurate species identification. Public health interventions, such as the use of protective equipment during high-risk activities and environmental management of drainage ditches, should be prioritized in the identified high-risk zones. This work demonstrates the critical importance of combining multiple diagnostic approaches to unravel the complex epidemiology of leptospirosis and provides a methodological framework for similar investigations in other endemic regions.
Notes
Competing interests
The authors declare that they have no competing interests.
References
[1] Perez J, Goarant C. Rapid Leptospira identification by direct sequencing of the diagnostic PCR products in New Caledonia. BMC Microbiol. 2010 Dec 22;10:325. DOI: 10.1186/1471-2180-10-325[2] Philip N, Bahtiar Affendy N, Ramli SNA, Arif M, Raja P, Nagandran E, Renganathan P, Taib NM, Masri SN, Yuhana MY, Than LTL, Seganathirajah M, Goarant C, Goris MGA, Sekawi Z, Neela VK. Leptospira interrogans and Leptospira kirschneri are the dominant Leptospira species causing human leptospirosis in Central Malaysia. PLoS Negl Trop Dis. 2020 Mar 23;14(3):e0008197. DOI: 10.1371/journal.pntd.0008197
[3] Boonsilp S, Thaipadungpanit J, Amornchai P, Wuthiekanun V, Bailey MS, Holden MT, Zhang C, Jiang X, Koizumi N, Taylor K, Galloway R, Hoffmaster AR, Craig S, Smythe LD, Hartskeerl RA, Day NP, Chantratita N, Feil EJ, Aanensen DM, Spratt BG, Peacock SJ. A single multilocus sequence typing (MLST) scheme for seven pathogenic Leptospira species. PLoS Negl Trop Dis. 2013;7(1):e1954. DOI: 10.1371/journal.pntd.0001954
[4] Guernier V, Richard V, Nhan T, Rouault E, Tessier A, Musso D. Leptospira diversity in animals and humans in Tahiti, French Polynesia. PLoS Negl Trop Dis. 2017 Jun 28;11(6):e0005676. DOI: 10.1371/journal.pntd.0005676
[5] Loureiro AP, Jaeger LH, Di Azevedo MIN, Miraglia F, Moreno LZ, Moreno AM, Pestana CP, Carvalho-Costa FA, Medeiros MA, Lilenbaum W. Molecular epidemiology of Leptospira noguchii reveals important insights into a One Health context. Transbound Emerg Dis. 2020 Jan;67(1):276-83. DOI: 10.1111/tbed.13349
[6] Piredda I, Ponti MN, Palmas B, Noworol M, Pedditzi A, Rebechesu L, Chisu V. Molecular Typing of Pathogenic Leptospira Species Isolated from Wild Mammal Reservoirs in Sardinia. Animals (Basel). 2021 Apr 13;11(4):1109. DOI: 10.3390/ani11041109
[7] Gajdov V, Jokic G, Savic S, Zekic M, Blazic T, Rajkovic M, Petrovic T. Genotyping of Leptospira spp. in wild rats leads to first time detection of L. kirshneri serovar Mozdok in Serbia. Front Microbiol. 2024 Mar 28;15:1379021. DOI: 10.3389/fmicb.2024.1379021
[8] Matthias MA, Díaz MM, Campos KJ, Calderon M, Willig MR, Pacheco V, Gotuzzo E, Gilman RH, Vinetz JM. Diversity of bat-associated Leptospira in the Peruvian Amazon inferred by bayesian phylogenetic analysis of 16S ribosomal DNA sequences. Am J Trop Med Hyg. 2005 Nov;73(5):964-74.
[9] Stoddard RA, Gee JE, Wilkins PP, McCaustland K, Hoffmaster AR. Detection of pathogenic Leptospira spp. through TaqMan polymerase chain reaction targeting the LipL32 gene. Diagn Microbiol Infect Dis. 2009 Jul;64(3):247-55. DOI: 10.1016/j.diagmicrobio.2009.03.014
[10] Cosson JF, Picardeau M, Mielcarek M, Tatard C, Chaval Y, Suputtamongkol Y, Buchy P, Jittapalapong S, Herbreteau V, Morand S. Epidemiology of leptospira transmitted by rodents in southeast Asia. PLoS Negl Trop Dis. 2014 Jun 5;8(6):e2902. DOI: 10.1371/journal.pntd.0002902
[11] Guernier V, Lagadec E, Cordonin C, Le Minter G, Gomard Y, Pagès F, Jaffar-Bandjee MC, Michault A, Tortosa P, Dellagi K. Human Leptospirosis on Reunion Island, Indian Ocean: Are Rodents the (Only) Ones to Blame? PLoS Negl Trop Dis. 2016 Jun 13;10(6):e0004733. DOI: 10.1371/journal.pntd.0004733
[12] Mason MR, Encina C, Sreevatsan S, Muñoz-Zanzi C. Distribution and Diversity of Pathogenic Leptospira Species in Peri-domestic Surface Waters from South Central Chile. PLoS Negl Trop Dis. 2016 Aug 16;10(8):e0004895. DOI: 10.1371/journal.pntd.0004895
[13] Kurucz K, Madai M, Bali D, Hederics D, Horváth G, Kemenesi G, Jakab F. Parallel Survey of Two Widespread Renal Syndrome-Causing Zoonoses: Leptospira spp. and Hantavirus in Urban Environment, Hungary. Vector Borne Zoonotic Dis. 2018 Apr;18(4):200-5. DOI: 10.1089/vbz.2017.2204
[14] Garcia-Lopez M, Lurier T, Bouilloud M, Pradel J, Tatard C, Sepulveda D, Anfray G, Dussert J, Bourhy P, Charbonnel N, Djelouadji Z. Prevalence, genetic diversity and eco-epidemiology of pathogenic Leptospira species in small mammal communities in urban parks Lyon city, France. PLoS One. 2024 Apr 10;19(4):e0300523. DOI: 10.1371/journal.pone.0300523
[15] Dobigny G, Garba M, Tatard C, Loiseau A, Galan M, Kadaouré I, Rossi JP, Picardeau M, Bertherat E. Urban Market Gardening and Rodent-Borne Pathogenic Leptospira in Arid Zones: A Case Study in Niamey, Niger. PLoS Negl Trop Dis. 2015 Oct 5;9(10):e0004097. DOI: 10.1371/journal.pntd.0004097
[16] Allan KJ, Biggs HM, Halliday JE, Kazwala RR, Maro VP, Cleaveland S, Crump JA. Epidemiology of Leptospirosis in Africa: A Systematic Review of a Neglected Zoonosis and a Paradigm for 'One Health' in Africa. PLoS Negl Trop Dis. 2015 Sep 14;9(9):e0003899. DOI: 10.1371/journal.pntd.0003899
[17] Sluydts V, Sarathchandra SR, Piscitelli AP, Van Houtte N, Gryseels S, Mayer-Scholl A, Bier NS, Htwe NM, Jacob J. Ecology and distribution of Leptospira spp., reservoir hosts and environmental interaction in Sri Lanka, with identification of a new strain. PLoS Negl Trop Dis. 2022 Sep 16;16(9):e0010757. DOI: 10.1371/journal.pntd.0010757
[18] Pricemou S, Soropogui B, Bérété F, Beavogui MB, Samoura A, Picardeau M, Bourhy P, Tordo N, Grayo S. Diversity of Leptospira Species and Their Rodent Reservoirs in the Guinean Forest. Microorganisms. 2025 Apr 7;13(4):833. DOI: 10.3390/microorganisms13040833
[19] Houéménou G, Gauthier P, Etougbétché J, Badou S, Dossou HJ, Agossou A, Picardeau M, Dobigny G. Pathogenic Leptospira in commensal small mammals from the extensively urbanized coastal Benin. Urban Sci. 2019 Sep 6;3(3):99. DOI: 10.3390/urbansci3030099
[20] Tangkanakul W, Tharmaphornpil P, Plikaytis BD, Bragg S, Poonsuksombat D, Choomkasien P, Kingnate D, Ashford DA. Risk factors associated with leptospirosis in northeastern Thailand, 1998. Am J Trop Med Hyg. 2000 Sep-Oct;63(3-4):204-8. DOI: 10.4269/ajtmh.2000.63.204
[21] Chadsuthi S, Modchang C, Lenbury Y, Iamsirithaworn S, Triampo W. Modeling seasonal leptospirosis transmission and its association with rainfall and temperature in Thailand using time-series and ARIMAX analyses. Asian Pac J Trop Med. 2012 Jul;5(7):539-46. DOI: 10.1016/S1995-7645(12)60095-9
[22] Suwanpakdee S, Kaewkungwal J, White LJ, Asensio N, Ratanakorn P, Singhasivanon P, Day NP, Pan-Ngum W. Spatio-temporal patterns of leptospirosis in Thailand: is flooding a risk factor? Epidemiol Infect. 2015 Jul;143(10):2106-15. DOI: 10.1017/S0950268815000205
[23] Mwachui MA, Crump L, Hartskeerl R, Zinsstag J, Hattendorf J. Environmental and Behavioural Determinants of Leptospirosis Transmission: A Systematic Review. PLoS Negl Trop Dis. 2015 Sep 17;9(9):e0003843. DOI: 10.1371/journal.pntd.0003843
[24] Crump JA, Mogeni P, Ajanovic SA, Bramugy JM, Chimenya M, Green EW, Lal S, Mabey DCW, Mayxay M, Newton PN, Olaru ID, Hopkins H, Picardeau M; Febrile Illness Evaluation in a Broad Range of Endemicities (FIEBRE) Consortium. Leptospirosis Prevalence and Risk Factors Among Patients Presenting With Fever to 4 Healthcare Sites in Sub-Saharan Africa and South East Asia: An International Multisite Observational and Nested Case-Control Study. J Infect Dis. 2026 Jan 17;233(1):e259-e270. DOI: 10.1093/infdis/jiaf464
[25] Kendall EA, LaRocque RC, Bui DM, Galloway R, Ari MD, Goswami D, Breiman RF, Luby S, Brooks WA. Leptospirosis as a cause of fever in urban Bangladesh. Am J Trop Med Hyg. 2010 Jun;82(6):1127-30. DOI: 10.4269/ajtmh.2010.09-0574
[26] Casanovas-Massana A, Pedra GG, Wunder EA Jr, Diggle PJ, Begon M, Ko AI. Quantification of Leptospira interrogans Survival in Soil and Water Microcosms. Appl Environ Microbiol. 2018 Jun 18;84(13):e00507-18. DOI: 10.1128/AEM.00507-18
[27] Costa F, Wunder EA Jr, De Oliveira D, Bisht V, Rodrigues G, Reis MG, Ko AI, Begon M, Childs JE. Patterns in Leptospira Shedding in Norway Rats (Rattus norvegicus) from Brazilian Slum Communities at High Risk of Disease Transmission. PLoS Negl Trop Dis. 2015 Jun 5;9(6):e0003819. DOI: 10.1371/journal.pntd.0003819
[28] Chirathaworn C, Inwattana R, Poovorawan Y, Suwancharoen D. Interpretation of microscopic agglutination test for leptospirosis diagnosis and seroprevalence. Asian Pac J Trop Biomed. 2014 May;4(Suppl 1):S162-4. DOI: 10.12980/APJTB.4.2014C580
[29] Muller SK, Assenga JA, Matemba LE, Misinzo G, Kazwala RR. Human leptospirosis in Tanzania: sequencing and phylogenetic analysis confirm that pathogenic Leptospira species circulate among agro-pastoralists living in Katavi-Rukwa ecosystem. BMC Infect Dis. 2016 Jun 10;16:273. DOI: 10.1186/s12879-016-1588-x
[30] Morey RE, Galloway RL, Bragg SL, Steigerwalt AG, Mayer LW, Levett PN. Species-specific identification of Leptospiraceae by 16S rRNA gene sequencing. J Clin Microbiol. 2006 Oct;44(10):3510-6. DOI: 10.1128/JCM.00670-06

